
Recommendation
Understanding factors explaining both intra and interspecific variation in susceptibility to infection by parasites remains a key question in evolutionary biology. Within a species variation in susceptibility is often explained by differences in behaviour affecting exposure to infection and/or resistance affecting the degree by which parasite growth is controlled (Roy & Kirchner, 2000, Behringer et al., 2000). This can vary between the sexes (Kelly et al., 2018) and may be explained by the ability of a parasite to attack different organs or tissues (Brierley et al., 2019). However, what goes on within one species is not always relevant to another, making it unclear when patterns can be scaled up and generalised across species. This is also important to understand when parasites may jump hosts, or identify species that may be susceptible to a host jump (Longdon et al., 2015). Phylogenetic distance between hosts is often an important factor explaining susceptibility to a particular parasite in plant and animal hosts (Gilbert & Webb, 2007, Faria et al., 2013).
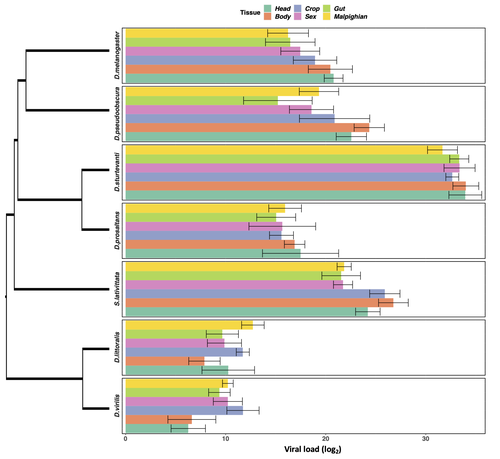
In two separate experiments, Roberts and Longdon (Roberts & Longdon, 2022) investigated how sex and tissue tropism affected variation in the load of Drosophila C Virus (DCV) across multiple Drosophila species. DCV load has been shown to correlate positively with mortality (Longdon et al., 2015). Overall, they found that load did not vary between the sexes; within a species males and females had similar DCV loads for 31 different species. There was some variation in levels of DCV growth in different tissue types, but these too were consistent across males for 7 species of Drosophila. Instead, in both experiments, host phylogeny or interspecific variation, explained differences in DCV load with some species being more infected than others.
This study is neat in that it incorporates and explores simultaneously both intra and interspecific variation in infection-related life-history traits which is not often done (but see (Longdon et al., 2015, Imrie et al., 2021, Longdon et al., 2011, Johnson et al., 2012). Indeed, most studies to date explore either inter-specific differences in susceptibility to a parasite (it can or can’t infect a given species) (Davies & Pedersen, 2008, Pfenning-Butterworth et al., 2021) or intra-specific variability in infection-related traits (infectivity, resistance etc.) due to factors such as sex, genotype and environment (Vale et al., 2008, Lambrechts et al., 2006). This work thus advances on previous studies, while at the same time showing that sex differences in parasite load are not necessarily pervasive.
References
Behringer DC, Butler MJ, Shields JD (2006) Avoidance of disease by social lobsters. Nature, 441, 421–421. https://doi.org/10.1038/441421a
Brierley L, Pedersen AB, Woolhouse MEJ (2019) Tissue tropism and transmission ecology predict virulence of human RNA viruses. PLOS Biology, 17, e3000206. https://doi.org/10.1371/journal.pbio.3000206
Davies TJ, Pedersen AB (2008) Phylogeny and geography predict pathogen community similarity in wild primates and humans. Proceedings of the Royal Society B: Biological Sciences, 275, 1695–1701. https://doi.org/10.1098/rspb.2008.0284
Faria NR, Suchard MA, Rambaut A, Streicker DG, Lemey P (2013) Simultaneously reconstructing viral cross-species transmission history and identifying the underlying constraints. Philosophical Transactions of the Royal Society B: Biological Sciences, 368, 20120196. https://doi.org/10.1098/rstb.2012.0196
Gilbert GS, Webb CO (2007) Phylogenetic signal in plant pathogen–host range. Proceedings of the National Academy of Sciences, 104, 4979–4983. https://doi.org/10.1073/pnas.0607968104
Imrie RM, Roberts KE, Longdon B (2021) Between virus correlations in the outcome of infection across host species: Evidence of virus by host species interactions. Evolution Letters, 5, 472–483. https://doi.org/10.1002/evl3.247
Johnson PTJ, Rohr JR, Hoverman JT, Kellermanns E, Bowerman J, Lunde KB (2012) Living fast and dying of infection: host life history drives interspecific variation in infection and disease risk. Ecology Letters, 15, 235–242. https://doi.org/10.1111/j.1461-0248.2011.01730.x
Kelly CD, Stoehr AM, Nunn C, Smyth KN, Prokop ZM (2018) Sexual dimorphism in immunity across animals: a meta-analysis. Ecology Letters, 21, 1885–1894. https://doi.org/10.1111/ele.13164
Lambrechts L, Chavatte J-M, Snounou G, Koella JC (2006) Environmental influence on the genetic basis of mosquito resistance to malaria parasites. Proceedings of the Royal Society B: Biological Sciences, 273, 1501–1506. https://doi.org/10.1098/rspb.2006.3483
Longdon B, Hadfield JD, Day JP, Smith SCL, McGonigle JE, Cogni R, Cao C, Jiggins FM (2015) The Causes and Consequences of Changes in Virulence following Pathogen Host Shifts. PLOS Pathogens, 11, e1004728. https://doi.org/10.1371/journal.ppat.1004728
Longdon B, Hadfield JD, Webster CL, Obbard DJ, Jiggins FM (2011) Host Phylogeny Determines Viral Persistence and Replication in Novel Hosts. PLOS Pathogens, 7, e1002260. https://doi.org/10.1371/journal.ppat.1002260
Pfenning-Butterworth AC, Davies TJ, Cressler CE (2021) Identifying co-phylogenetic hotspots for zoonotic disease. Philosophical Transactions of the Royal Society B: Biological Sciences, 376, 20200363. https://doi.org/10.1098/rstb.2020.0363
Roberts KE, Longdon B (2023) Heterogeneities in infection outcomes across species: examining sex and tissue differences in virus susceptibility. bioRxiv 2022.11.01.514663, ver. 2 peer-reviewed and recommended by Peer Community in Evolutionary Biology. https://doi.org/10.1101/2022.11.01.514663
Roy BA, Kirchner JW (2000) Evolutionary Dynamics of Pathogen Resistance and Tolerance. Evolution, 54, 51–63. https://doi.org/10.1111/j.0014-3820.2000.tb00007.x
Vale PF, Stjernman M, Little TJ (2008) Temperature-dependent costs of parasitism and maintenance of polymorphism under genotype-by-environment interactions. Journal of Evolutionary Biology, 21, 1418–1427. https://doi.org/10.1111/j.1420-9101.2008.01555.x
Authors replied to all comments, and carefully addressed all suggestions. Again, I really like the manuscript. I think the revision significantly improved the paper. I recommend publication without more comments. Congratulations for this nice work.
https://doi.org/10.24072/pci.evolbiol.100638.rev21This paper has been revised well in line with my comments and is now ready for publication.
https://doi.org/10.24072/pci.evolbiol.100638.rev22DOI or URL of the preprint: https://doi.org/10.1101/2022.11.01.514663
Version of the preprint: 1
Please find our response attached.
I agree with both reviewers that this preprint is interesting, well written and will make a good contribution to PCI Evolutionary Biology. The preprint combines 2 experiments addressing sex differences in susceptibility to DCV across 31 different species and tissue tropism across males for 7 species. As it stands, the preprint is of high quality, however both reviewers make some useful comments. Reviewer 1 suggests that framing the preprint in terms of ‘heterogeneity in infection outcomes’ that might arise due to sex or tissue tropism may make the flow of the introduction a bit easier. Both reviewers also suggest that additional information about sex differences in infection may add to the preprint -notably Reviewer 1 suggests more information about STIs and how this differs to ordinary infectious diseases and Reviewer 2 some text explaining why sex differences in infections might arise due to a trade-off with reproduction. There are a few typos that I highlighted in the attached pdf.
Download recommender's annotationsRoberts and Longdon
This is an interesting and thorough paper on sex differences in viral progression across drosophilids, and repeatability of tropism patterns. I found the methods and analysis well described (though note slightly above my expertise in analysis here) and the results clear.
I've attached a pdf with comments on for minor style issues. I'd ask the authors to consider the following:
a) There is a body of work on sex biased infection from the STI literature (Lockhart Biol Rev. is an entry point,also Knell and Webberley and some others), which does tend to find sex biased infection (through exposure differences e.g Ryder et al 2014 Am Nat) and sex biased virulence patterns (e.g. female limited sterility or female biased sterility). This literature also goes across to insects, so I would suggest mentioning this as a separate case and then examining what are sometimes (after Lockhart) known as Ordinary Infectious Diseases.
b) The piece tries to do two things in one paper, which is always a tricky ask in terms of writing. It's not needed, but does make the introduction a little saltatory. I wonder if an overall framing in terms of 'heterogeneity in infection outcome' might be a way forward, saying this can be viewed both at the level of individuals and species.E.g sex of host wihtin species, pattern of infection between species .
c) The start of the discussion would benefit from a brief recapitulaiton of the main aims/ideas before reconciling to the literature.
d) Whilst experiments and analyses were strong to me, I think an important caveat for the discussion is that the experiments on sex bias were limited to i) young flies and ii) largely virgin flies. I would add 'exploration in a few systems should investigate if age and matedness status alter these results'. Further, the tropism were limited to a sex and age as well, so again a caution that tropism did not vary in young male flies is the formal conclusion, rather than there is no variation in tropism amongst species (which is true of generalised).
Aside these, minor comments only. I enjoyed this paper and it clearly has a substantial body of work that can be published - just a matter of polish and the occasional tempering caveat.
Download the review https://doi.org/10.24072/pci.evolbiol.100638.rev11
The preprint “Sex and tissue differences in virus susceptibility across species” reports the results of two experiments where Drosophila C Virus (DCV) genomes has been quantified by quantitative RT-PCR in males and females (experiment 1: effect of sex) and different tissues (experiment 2) of several species of fruit flies of the family Drosophilidae.
The first experiment was performed over 31 species, and showed no clear difference in viral loads between sexes. Although viral load varied greatly between species, males and females viral load were rather consistent between sexes of the same species, with a close to 1:1 overall ratio between sexes. Consistent with a previous paper of the same authors, host phylogeny explained most of the observed interspecific variation. Surprisingly, there was no effect of body size on viral, which I found particularly interesting, both within species and across species. Authors also found no effect of mating status, but I believe that the absence of effect is largely due to the experiment not designed to test it (only 4 species showed sign of uncontrolled mating).
The second experiment was performed over 7 species of flies, where males (only) were infected and dissected to measure viral load in different body parts/tissues. The main result here is again a strong effect of species on viral loads, and very little within species variation and across species tissue consistency. This is a very interesting study that investigated the possible effect of tissue tropism to explained variation in disease virulence across species, or interspecific variation of disease transmission.
Overall, I found this paper well written and very clear. The rational of the study is clear, the experiments are sound, without apparent flaw, and their analysis clearly described. I really enjoyed reading this preprint and I believe it provides highly valuable results and was very stimulating. I do not have comments to make on the experimental procedures, as I think experiments were done properly and reported clearly. I will only make comments on the report itself, with a few suggestions which I hope will be helpful to authors.
Major suggestions:
In the second paragraph of the introduction, I found the background on sex differences in immunity in invertebrates a little light, with no mention of reproduction trade-off theory that may explain why males are generally more susceptible to diseases than females. One important paper to cite may be about the Bateman’s principle in insect immunity (https://doi.org/10.1098/rspb.2002.1959).
It is unclear to me why authors decided to report statistical significance from experiment 1 based on CI, and p-value for experiment 2. Is this because of different statistical models used? It may be better to announce the level of significance for each model in the method section.
Why not illustrating the phylogenetic effects and the absence of effect of body size in experiment 1, even in a supplementary figure? These are valuable results, and I would have welcomed more plots.
Minor comments:
Line 23: I would add for clarity in the abstract “to DCV”, in the sentence “suggesting that susceptibility [to DCV] is not sex specific”. Otherwise, it sounds like a general statement.
Line 55: missing letter, please correct “parasitism rates in mammals”
When citing species names, please add a space between genus initial letter and species name, throughout the paper.
Line 132, authors justified that pricking flies with DCV does not change the overall distribution of viruses across fly tissue compared to oral feeding. But authors may acknowledge this might be true for D. melanogaster only (as cited in ref. 28). I am not aware of other study testing this, and based on the rational of the present ms, one cannot rule out a transmission route by species interaction leading to a difference in tissue tropism.
In the section ‘Inferring the host phylogeny’, not all references are cited as number. Citation format should be homogenized.
Line 233 and 251: It is unclear to me what is strain-specific effect and variance. Is this fly species? Please clarify.
Line 335-336: I think this sentence missed its reference (meta-analysis).
https://doi.org/10.24072/pci.evolbiol.100638.rev12
