
Phenotypic stasis despite genetic divergence and differentiation in Caenorhabditis elegans.
Phenotypic stasis with genetic divergence
Abstract
Recommendation: posted 17 November 2023, validated 20 November 2023
Guillaume, F. (2023) Phenotypic stasis despite genetic divergence and differentiation in Caenorhabditis elegans.. Peer Community in Evolutionary Biology, 100627. https://doi.org/10.24072/pci.evolbiol.100627
Recommendation
Explaining long periods of evolutionary stasis, the absence of change in trait means over geological times, despite the existence of abundant genetic variation in most traits has challenged evolutionary theory since Darwin's theory of evolution by gradual modification (Estes & Arnold 2007). Stasis observed in contemporary populations is even more daunting since ample genetic variation is usually coupled with the detection of selection differentials (Kruuk et al. 2002, Morrissey et al. 2010). Moreover, rapid adaptation to environmental changes in contemporary populations, fuelled by standing genetic variation provides evidence that populations can quickly respond to an adaptive challenge. Explanations for evolutionary stasis usually invoke stabilizing selection as a main actor, whereby optimal trait values remain roughly constant over long periods of time despite small-scale environmental fluctuations. Genetic correlation among traits may also play a significant role in constraining evolutionary changes over long timescales (Schluter 1996). Yet, genetic constraints are rarely so strong as to completely annihilate genetic changes, and they may evolve. Patterns of genetic correlations among traits, as captured in estimates of the G-matrix of additive genetic co-variation, are subject to changes over generations under the action of drift, migration, or selection, among other causes (Arnold et al. 2008). Therefore, under the assumption of stabilizing selection on a set of traits, phenotypic stasis and genetic divergence in patterns of trait correlations may both be observed when selection on trait correlations is weak relative to its effect on trait means.
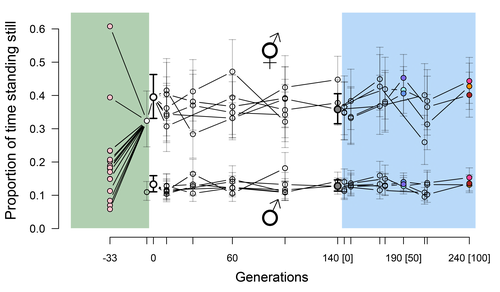
Mallard et al. (2023) set out to test whether selection or drift may explain the divergence in genetic correlation among traits in experimental lines of the nematode Caenorhabditis elegans and whether stabilizing selection may be a driver of phenotypic stasis. To do so, they analyzed the evolution of locomotion behavior traits over 100 generations of lab evolution in a constant and homogeneous environment after 140 generations of domestication from a largely differentiated set of founder populations. The locomotion traits were transition rates between movement states and direction (still, forward or backward movement). They could estimate the traits' broad-sense G-matrix in three populations at two generations (50 and 100), and in the ancestral mixed population. Similarly, they estimated the shape of the selection surface by regressing locomotion behavior on fertility. Armed with both G-matrix and surface estimates, they could test whether the G's orientation matched selection's orientation and whether changes in G were constrained by selection. They found stasis in trait mean over 100 generations but divergence in the amount and orientation of the genetic variation of the traits relative to the ancestral population. The selected populations changed orientation of their G-matrices and lost genetic variation during the experiment in agreement with a model of genetic drift on quantitative traits. Their estimates of selection also point to mostly stabilizing selection on trait combinations with weak evidence of disruptive selection, suggesting a saddle-shaped selection surface. The evolutionary responses of the experimental populations were mostly consistent with small differentiation in the shape of G-matrices during the 100 generations of stabilizing selection.
Mallard et al. (2023) conclude that phenotypic stasis was maintained by stabilizing selection and drift in their experiment. They argue that their findings are consistent with a "table-top mountain" model of stabilizing selection, whereby the population is allowed some wiggle room around the trait optimum, leaving space for random fluctuations of trait variation, and especially trait co-variation. The model is an interesting solution that might explain how stasis can be maintained over contemporary times while allowing for random differentiation of trait genetic co-variation. Whether such differentiation can then lead to future evolutionary divergence once replicated populations adapt to a new environment is an interesting idea to follow.
References
Arnold, S. J., Bürger, R., Hohenlohe, P. A., Ajie, B. C. and Jones, A. G. 2008. Understanding the evolution and stability of the G-matrix. Evolution 62(10): 2451-2461.
https://doi.org/10.1111/j.1558-5646.2008.00472.x
Estes, S. and Arnold, S. J. 2007. Resolving the Paradox of Stasis: Models with Stabilizing Selection Explain Evolutionary Divergence on All Timescales.. Am. Nat. 169(2): 227-244.
https://doi.org/10.1086/510633
Kruuk, L. E. B., Slate, J., Pemberton, J. M., Brotherstone, S., Guinness, F. and Clutton-Brock, T. 2002. Antler size in red deer: Heritability and selection but no evolution. Evolution 56(8): 1683-1695.
https://doi.org/10.1111/j.0014-3820.2002.tb01480.x
Mallard, F., Noble, L., Guzella, T., Afonso, B., Baer, C. F., Teotónio, H. 2023. Phenotypic stasis with genetic divergence. bioRxiv, ver. 3 peer-reviewed and recommended by Peer Community in Evolutionary Biology.
https://doi.org/10.1101/2022.05.28.493856
Morrissey, M. B., Kruuk, L. E. B. and Wilson, A. J. 2010. The danger of applying the breeder's equation in observational studies of natural populations. J Evolution Biol 23(11): 2277-2288.
https://doi.org/10.1111/j.1420-9101.2010.02084.x
Schluter, D. 1996. Adaptive radiation along genetic lines of least resistance. Evolution 50(5): 1766-1774.
https://doi.org/10.1111/j.1558-5646.1996.tb03563.x

The recommender in charge of the evaluation of the article and the reviewers declared that they have no conflict of interest (as defined in the code of conduct of PCI) with the authors or with the content of the article. The authors declared that they comply with the PCI rule of having no financial conflicts of interest in relation to the content of the article.
This work was supported by the European Research Council (ERC-St-243285) and the Agence Na- tionale pour la Recherche (ANR-14-ACHN-0032-01, ANR-17-CE02-0017-01) to HT, the National Institutes of Health (R01GM107227) to CB, and a Marie Curie fellowship (H2020-MSCA-IF-2017- 798083) to LN. This research has also received support, under the KITP Quantitative Biology program, from the National Science Foundation (PHY-1748958) and from the Gordon and Betty Moore Foundation (2919.02).
Evaluation round #2
DOI or URL of the preprint: https://doi.org/10.1101/2022.05.28.493856
Version of the preprint: 2
Author's Reply, 31 Oct 2023
Paris, 31 October 31, 2023
Dear Frédéric,
I am re-submitting the manuscript “Phenotypic stasis with genetic divergence” for your recommendation to PCI Evolutionary Biology. It should come up shorty in bioRxiv (doi: https://doi.org/10.1101/2022.05.28.493856)
We thank you for the comments and suggestions that you made. In the file attached you will see our point-by-reply to your concerns. We believe to have addressed them by performing new analyses, and clarifying some of our arguments, though we have not been able to follow all of your suggestions.
Again, we appreciate the time and effort you (and the referees) put in reviewing our work. We believe that the manuscript has been much improved and hope that you will find it appropriate for PCI Evolutionary Biology.
On behalf of the authors,
Henrique
Decision by Frédéric Guillaume , posted 18 Aug 2023, validated 18 Aug 2023
, posted 18 Aug 2023, validated 18 Aug 2023
Thank you for the comprehensive revisions of your manuscript which addresses the main points raised by the two previous reviewers and myself. After reevaluation of your work, I found a few more points to address and clarify before I can write a recommendation.
Overall, if writing a recommendation from the current manuscript, I would summarize your findings as being in line with a model of stabilizing selection with drift on a set of locomotion traits. The main evolution of the traits during stasis is a general loss of genetic variance across the board, which is compatible with a model of stabilizing selection after hybridization among divergent inbred lines, but also with drift. There is no need to invoke directional selection at this stage (see below). The transient changes in G-matrix orientation seem rather limited, although not fully explored, limiting our ability to draw strong conclusions on their importance. Similarly, the relative importance of selection and drift has not been formally tested. The study thus remains a general assessment of changes in genetic variation over time without a strong conclusion regarding the causes and consequences of the changes.
I think you can strengthen your manuscript if you address the points raised here, especially regarding the changes in G and the selection regime.
Main comments:
- Directional selection: I am still not convinced that you have a strong case for directional selection in the domestication phase (from A0 to A6140), in contrast to your argumentation for it, eg. on lines 529-531, 558-559. As point in case, you provide ample evidence of a lack of directional change in the traits during that period. It is thus misleading to invoke directional selection in the Discussion to explain genetic divergence.
I am not denying that there might have been directional selection during hybridization of the founders, but apparently there was no directional selection during domestication, and it is not necessary to explain your results. The manuscript should clarify this point.
Your results can be interpreted as stemming from the effects of stabilizing selection on an initially broad phenotypic/genotypic distribution (A0) generated by admixture of the inbred founder lines and thus large segregation variance. Stabilizing selection sifts through that initial variance, eliminating extreme phenotypes during domestication and eventually leading to a selection-mutation-drift-recombination equilibrium. There is no need here to consider directional selection on the traits nor "effective" stabilizing selection, just stabilizing selection is enough. Please make this clearer in the text (ie, lines 529-531, 533-535, 558-559).
Also, the model of stabilizing selection on a plateau and directional selection outside the plateau that you mention is, in my opinion, not different from a model of stabilizing selection with a broad adaptive surface around the optimum (at least broad relative to the phenotypic variance). This doesn't deny directional selection when the population mean is away from the optimum. This is to clarify my thoughts, not to ask you to change your phrasing but you can make the parallel with the classical model of stabilizing selection for a multi-trait optimum with a broad selection surface. - G-matrix divergence and orientation: divergence in G-matrix orientation is not directly evaluated in the Results section but mentioned in the Discussion (lines: 557, 648), and also regarding alignment with the selection surface (lines 591-593). Most mentions of alignments are relative to gmax of A6140. You do not explicitly test for a change in orientation of gmax in the CA populations. Assessment of changes in CA lines can bring evidence of transient changes in genetic covariances and arguments in favor of their importance relative to selection. In particular, it would help asses if "cryptic evolution of genetic covatiance" has happened (line 648). I have two requests:
- clarify alignment of G's between A6140 & CA-100; reduced variance along one phenotypic dimension can be caused by the general reduction in mean variance (smaller trace) and changes in orientation of G. As of now, it is not clear what are the changes in G's orientation in the CA lines and how they affect their reduced variation along gmax of A6140. Interpretation of the tensor analysis relative to changes in orientation of G's is not clear to me. A more direct comparison of G's alignement will help you clarify the changes in alignment.
- clarify alignment of selection surface with G's of CA lines; similarly, it is not clear how changes in G's orientation in CA lines affect patterns of genetic variance along axes of the selection surface nor how G-matrices align with that surface. Only mention is about alignment of y3 and gmax of A6140 (lines 523-524). A direct comparison with G's orientation in the CA lines is also warranted.
- Testing for differentiation by drift vs. selection: It might be that drift is the most important driver after domestication, but we do not know the relative importance of stabilizing selection vs. drift because you do not formally test for it. Also, it is unclear how you tested for a loss of variance matching with drift expectations. Please provide the model you are using and how you parameterized it. It is unclear how you modeled the multivariate loss of variance along axes of the selection surface with that model.
This calls for a multivariate Fst-Qst analysis to test whether genetic changes are dominated by drift or selection, over time or between lines. My understanding is that you have genomic data for some of the lines, from which you could estimate an Fst distribution. That additional analysis is likely demanding but would greatly add to the understanding of the system. A suggested approach is driftsel by Ovaskainen et al. (A new method to uncover signatures of divergent and stabilizing selection in quantitative traits. 2011. Genetics 189(2): 621-632; Karhunen et al. 2013. driftsel: an R package for detecting signals of natural selection in quantitative traits. Mol Ecol Resour 13(4): 746-754). - QTL data (lines 567-569): this mention of two QTL results brings more confusion than clarity. It is unclear how only two QTL of unknown effect size could be responsible for the maintenance of genetic variation in the locomotion traits. What about genome-wide mutations over 140 generations in a population with N=10^4? That part could be removed.
Additional comments:
- Divergence versus differentiation: I understand that you use the two words to differentiate the "between" and "within" generation differences in G's, while using the same method. This is confusing. Both words bear different meanings and would normally be tested with different methods. Differentiation is usually assessed with a Qst index (or Fst), for instance (ie, a ratio of variance). I would clarify the use you make of the two terms relative to your two types of comparison, or find other words.
- Abstract: lines 14-16, it is unclear what are you referring to with "large phenotypic scale" and "local phenotypic scale", no mention of such scales are done in the rest of the manuscript.
Corrections:
l55 : "little unexplored" -> "little explored", or "unexplored"
l119: please specify which population is used for the samples
l124: "generation 4-6 generations" -> phrasing unclear
l524: "Overall there is a strong alignment of both divergence and differentiation axes with y3 (Figure S12), and thus with gmax (see above)" -> phrasing unclear, it means that "divergence and differentiation axes" are aligned with gmax, but I guess you mean to say that y3 is aligned with gmax. Also clarify where exactly we should be looking "above".
l573: "however, the strength selection" -> "however, that the strength of selection"
Figure 5: please add the meaning of the color code as a legend
Evaluation round #1
DOI or URL of the preprint: https://doi.org/10.1101/2022.05.28.493856
Author's Reply, 07 Jul 2023
Decision by Frédéric Guillaume, posted 14 Dec 2022, validated 14 Dec 2022
Thank you for submitting this very interesting manuscript to PCI Evolutionary Biology. I have received two reviews of your manuscript. Both find interest in publishing your findings, with one very enthusiastic reviewer. I do also find value in the reported data and analyses. There are nevertheless some major points that have been raised by both reviewers, and myself, which need to be addressed in a revision of your work. The suggested revisions will necessitate some complementary analyses on the effect of sampling of the inbred lines in the CA populations and some additional effort with streamlining and focusing of the text, especially in the Results and Discussion sections.
I would like you to pay particular attention to the comments of reviewer 2. I do also find that the marked difference in G-matrix size between A6140 and CA1-3 are suspicious. Please carefully address the reviewer's point here. I'd suggest that you use the down-sampling of A6140 inbred lines to evaluate the effect of smaller number of inbred lines in CA1-3 on the reduction of the trace of G-matrices in CA1-3.
Similarly, the discussion is in part off topic when discussing un-observed increase in variance relative to ancestral population. I did not find any evidence reported in your manuscript about the increase of variance in y5-y6 as suggested in lines 496-498. In fact, reduction of variance along those axes of selection is reported previously, lines 475-476. The following discussion start with "If true" (line 498), speculation that doesn't seem supported by your data.
The first reviewer points to the lack of a better discussion on the implications of your study for the field. I would add to this that the manuscript is in general rather dry and technically oriented which will be a deterrent for a more generalist audience. I would strongly suggest to bring in more biological examples or interpretation of some of the methods, especially those dealing with canonical analyses of covariance matrices. Some basic mention of the biological meaning of vectors in phenotypic space should be added.
I would add that I do not find strong evidence for "squashed stabilizing selection" in your manuscript as mentioned on lines 469-470. At best, it suggests such possibility, but my understanding is that squashed stabilizing selection requires some form of frequency-dependent selection. More analysis or justification for a statement about whether your finding "provide evidence" of such selection regime are necessary.
Reviewed by Benoit Pujol, 17 Nov 2022
Review by Benoit Pujol for PCI Evol Biol of the preprint entitled “Phenotypic stasis with genetic divergence”, written by Mallard et al. (https://doi.org/10.1101/2022.05.28.493856; this version posted May 29, 2022.).
This is one of the most interesting paper I was given to read in a long time! I found it very clear. The authors did a brilliant job at presenting complex mechanisms (and the complex methods used to evaluate them) in a way that is easily understandable. Understanding how and why stasis can occur at mutation-selection equilibrium after a history of adaptation without blocking other evolutionary processes that shape the diversity of populations such as divergence is crucial. This is important because the disconnection between the theory and the empirical results found in wild populations is widely explained by the action of this or this mechanism. We need to integrate this perspective on mechanisms influencing the evolutionary potential of populations to respond to selection in a broader evolutionary perspective of stasis.
Below you will find comments, many of them positive, and some suggestions. I have only one point that I’d really like the authors to take into account (see my last comment): the implication of this work in terms of contribution to the knowledge in the domain and how it participates to push forward the theory is not clearly stated. I’d like to see it addressed by the authors.
Introduction
This is a particularly well written paper. Theoretical concepts and empirical evidence are elegantly reviewed and synthesized in the introduction before getting to the core of the proposed research. This presentation of the state of art illustrates a clear understanding of the complex topic of phenotypic stasis.
L30-31 I’m not a native English speaker and I trust your writing skills. Please check that this sentence is correct. I was in doubt because “important” came right after drift.
L42 These studies indicate… I totally agree with your point that a multivariate approach is necessary to understand stasis, and you have already mentioned that other mechanisms were at play (plasticity, indirect environmental covariance, etc.). Obviously, other mechanisms should be considered to and it’s impossible to investigate all of them simultaneously. It is only possible to study them in isolated studies, even in a given study system. If you feel like saying a word about this, feel free to do so.
L50 Aren’t we missing a “to” btw “expected” and “facilitate”
I really enjoyed reading your paragraph about selection in finite populations, that includes a perspective on drift
L 66 Aren’t we missing a comma after “occurred”
Ll 74-86 I was missing something here between the conceptual state of art and the down to earth presentation of the experiment. I think that moving the last sentence of this paragraph: “We seek to … … once most adaptation has been achieved.” At the beginning of the paragraph would help connecting these aspects.
Methods
I’m no big expert but I saw no flaw in the archived data and codes, and laboratory culture sections.
I am not able to evaluate the “Worm Tracking assays” section. It nevertheless seems fit in terms of using multiple generations, and sample replication for phenotypic measurements.
Ll173-174 Not sure to understand. Do you mean that they were not significant as covariables in a linear regression analysis and you therefore removed them before fitting an even fitter model? Or that you used the residuals of the linear regression as data to feed the model fitting procedure.
I think that the differentiation approach btw males and hermaphrodites is correct. I might have preferred other (more direct) approaches but I can understand why the authors did it this way in this study system.
The “locomotion behavior” section seems sound to me (notably the LMM approaches). I have to admit that I’m not a specialist of transition rates.
Ll226-229 “As we … … derived” Anyway, there is no “true” estimate of COVa since nobody can exclude unexpected indirect and/or environmental covariances, and you then explain that you estimate a broad-sense G matrix. You mention the assumptions but it is difficult to predict what bias it may generate in the results, if any. If you feel that it is justified, do not hesitate to add a few words about it.
L 259 please check whether “were fit with” is correct phrasing
L275 “this decomposition of describes” sounds like we’re missing a word here, please check
The “transition rate genetics” and “selection on transition rates” sections are clearly exposed the methods that will be used to estimate genetic covariances, compare G matrices and estimate their alignment with the selection surface.
The “inference of effects” section is also clearly presented and denotes objectivity in the analysis of effects. Taking into account the quantitative overlap between the credible intervals of expected distribution of random effects VS posterior distribution of effects is much more appropriate than model fit comparisons.
Results
The results section globally reads well, but I can’t help thinking that it includes a lot of methodological aspects that belong to the methods rather than the results in the sections 4.1, 4.2, 4.3. I appreciated that you remind the reader (me in particular) what is done because it is a dense paper that includes complex methods but it would be good if the focus was ore on the results than the methods in the method section of the main text. This is very minor and the take home message of my evaluation of the result section is that I greatly appreciated that the authors took great care to make their results accessible to a broad readership.
Ll 329-348 these elegant and easily understandable synthetic sentences about the lab culture read nicely. It nevertheless feels like I already read some of it in the methods and the Figure legend. Please check for direct redundancy.
There are many figures (when including the supplementary) to review, but I understand why they are necessary. On a global note, the legends are clear but the figure themselves are not always easy to read because text, bars, etc appear very small/thin.
Discussion
The discussion about the role of selection pressures and their signature on G matrices (with a full overview of the effect of divergent, stabilizing selection and drift) is very clear and elegant. I have to admit that I had to read the paper a few times to fully grasp all of it.
I appreciated very much the honesty of the authors who mentioned the various scenarii that could explain their results (original lack of standing variation, sampling error, etc.)
I agree that the indirect effects of environmental covariances, which could be interacting with this kind of study, is unlikely to underly unexpected genetic variance in this particular study. Pleiotropic effects are indeed a possible underlying mechanism, but I’d tend to say that this is not a wild population and that selection was artificially directed at these traits. I’m not saying that it excludes the potential pleiotropy but it will be a lot more messy in the wild. I find that you could have included more information about the balancing selection hypothesis. I find fluctuating selection is very interesting because its impact can vary from loss to gain in genetic variance depending on the intensity and frequency of directional changes, which generates very interesting processes. I’m not saying that you are in the presence of fluctuating selection. I’m merely saying that a bit more information about the features of the potential balancing selection at play here, in a context of stabilizing selection, would make this point more interesting and clear to me, and potentially others.
I wrote this before reading the next paragraph: “”About divergence, I wonder (since I’m not basing this idea on thorough knowledge) whether the interaction of random processes with a somewhat polygenic determinism may have unexpected consequences. I’m not saying that you should consider this idea in the text. I can’t fully wrap my head around it. I’m just putting this here, in case you know something about this that I don’t.”” It is therefore only logical that I say that I appreciated your discussion of the departure from the infinitesimal model.
Ll 591-593 These are indeed the most described mechanisms but many others are at play and were described in papers.
The conclusion, as well as the whole paper, is very clear. I find that the conclusion fails to present the fundamental implication of this brilliant study. It is necessary to clarify why and how the research presented here contributes to move the domain and the theory forward. In terms of natural history, it seems to me that the situation described here is potentially existing in many other wild populations that are described as being in stasis because they are at mutation-selection equilibrium post adaptation. I have my idea but I’d like you to state why it is important to show that genetic divergence can occur while there is a stasis.
https://doi.org/10.24072/pci.evolbiol.100627.rev11




